Life History Plasticity and Evolution
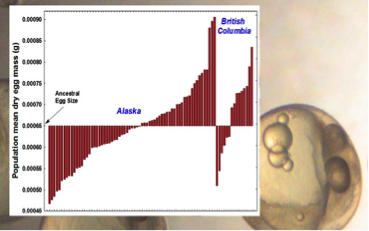
Phenotypic aspects that directly affect offspring production may be more intimately linked to fitness than any other organismal trait. In females, several of these aspects—including the age at which reproduction is initiated, the level of reproductive effort, and the size and number of offspring—collectively comprise one integrated and highly complex “trait”, the “reproductive life history”. The stickleback adaptive radiation in recently deglaciated regions, encompassing many thousands of independently evolving populations living in an incredibly diverse array of environments, is an especially useful model system for probing this evolution. Our research into life-history evolution, primarily in females, but also involving some aspects of male reproduction, takes place across a broad front. Our nearly 25-year set of collections from widely divergent habitats in Alaska, British Columbia, and more recently Newfoundland, permit an unparalleled view of how broadly life-history traits have evolved from the ancestral state, and in which direction—and to probe the environmental features underlying the divergence. A significant augmentation of this field research is made possible by our 325+ tank live fish room, which gives us the ability to rear hundreds of individuals of numerous populations each year, permitting us to perform experiments to test and extend our findings based on field collections. In most organisms, and in fish in particular, the strongest selection is upon the earliest life stages—in stickleback, their first month of life. We complement our field and laboratory work on adult life histories with fine-scale experiments on embryos, fry, and early juvenile stages of stickleback. These experiments allow us to infer the effects of factors such as starting egg size, food abundance, the level of competition, and even maternal size, condition, and degree of stress, on offspring growth and performance (Miguel Reyes, Melissa Graham). This holistic set of approaches has given us a unique opportunity to deeply probe the processes through which life histories evolve.
Some of our more intriguing findings include: a) all oceanic “ancestral” populations are not the same—a situation which raises the question of exactly what ancestor colonized a particular region (Richard King) b) the broad array of solutions stickleback populations have evolved to deal with invasion of novel freshwater environments—there are relatively few commonalities, with each population to some extent “finding its own path” to reproductive fitness; c) the rapidity with which life-history traits can evolve—we have detected some of the fastest phenotypic evolutionary rates that have been reported; and d) life-history traits vary tremendously in their degree of plasticity, and in the range of expression of any plasticity. Egg size shows no substantial plasticity within a single reproductive season, for example, but consistently shows plasticity across sequential ages. A general explanation for this finding is currently lacking.