The results presented here are based on:
A. Justo & D.S. Hibbett. 2011. Phylogenetic classification of Trametes (Basidiomycota, Polyporales) based on a five-marker dataset. Taxon 60(6): 1567-1583
Available online or by request to ajusto@clarku.edu
Abstract: The phylogeny of Trametes and related genera was studied using molecular data from ribosomal markers (nLSU, ITS) and protein-coding genes (RPB1, RPB2, TEF1-alpha) and consequences for the taxonomy and nomenclature of this group were considered. Separate datasets with rDNA data only, single datasets for each of the protein-coding genes and a combined five-marker dataset were analyzed. Molecular analyses recover a strongly supported trametoid clade that includes most of Trametes species (including the type T. suaveolens, the T. versicolor group, and mainly tropical species such as T. maxima and T. cubensis) together with species of Lenzites and Pycnoporus and Coriolopsis polyzona. Our data confirm the positions of Trametes cervina (= Trametopsis cervina) in the phlebioid clade and of Trametes trogii (= Coriolopsis trogii) outside the trametoid clade, closely related to Coriolopsis gallica. The genus Coriolopsis, as currently defined, is polyphyletic, with the type species as part of the trametoid clade and at least two additional lineages occurring in the core polyporoid clade. In view of these results the use of a single generic name (Trametes) for the trametoid clade is considered to be the best taxonomic and nomenclatural option as the morphological concept of Trametes would remain almost unchanged, few new nomenclatural combinations would be necessary and the classification of additional species (i.e. not yet described and/or sampled for molecular data) in Trametes based on morphological characters alone will still be possible. Alternative scenarios to divide the trametoid clade in five or ten genera were considered but if any of these options were to be adopted morphological distinction of the segregated genera and the ascription of additional species to any of these genera would be very difficult and in some cases impossible. The genera Artolenzites, Coriolopsis (as currently typified), Coriolus, Cubamyces, Cyclomycetella, Lenzites, Poronidulus, Pseudotrametes and Pycnoporus are considered synonyms of Trametes.

Introduction
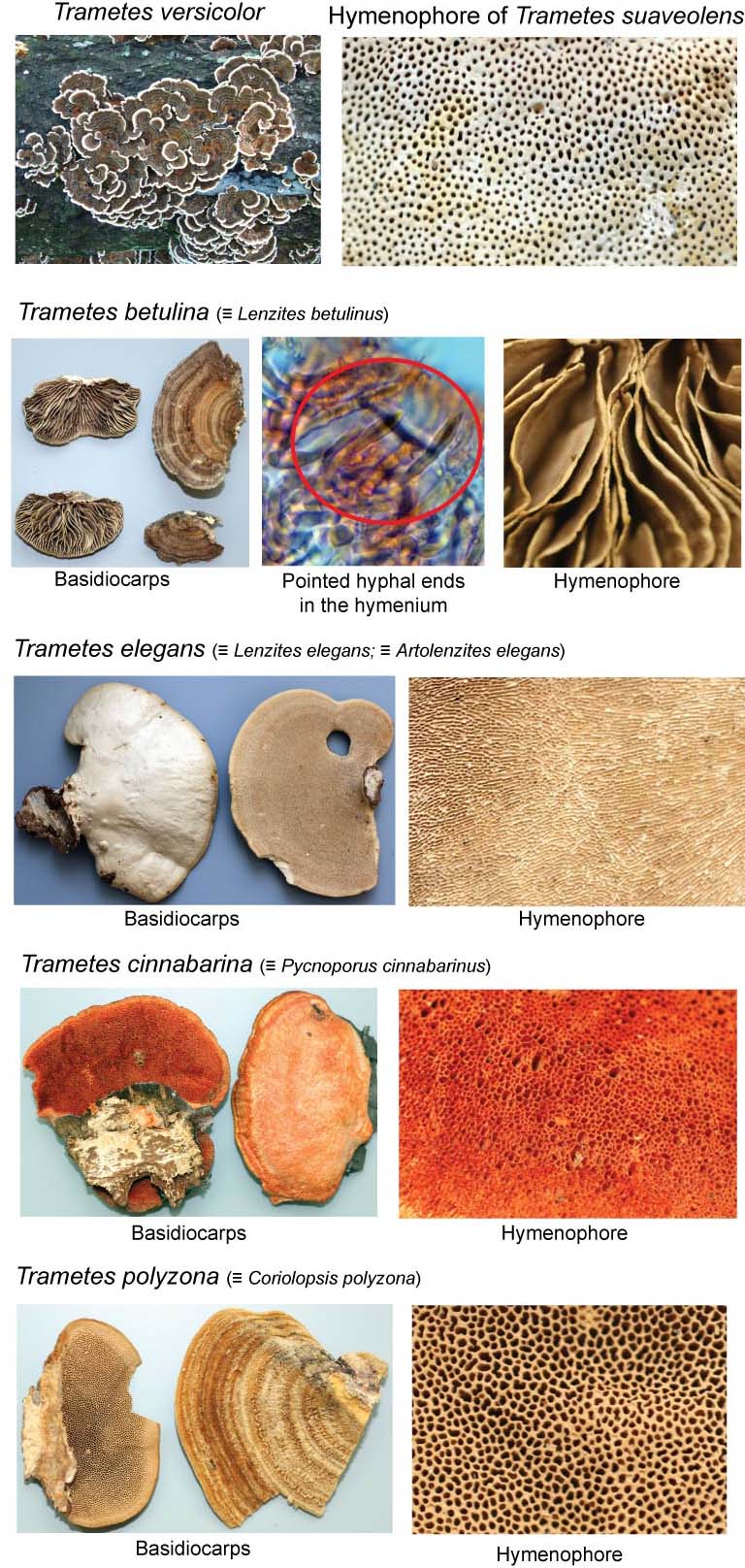
The genus Trametes Fr. (Polyporales, Basidiomycota) is characterized by the combination of pileate basidiocarps, poroid hymenophore, trimitic hyphal system, thin-walled smooth basidospores not reacting in presence of Melzer’s reagent and production of a white-rot type of wood decay (Gilbertson & Ryvarden, 1987). It is a cosmopolitan genus present in virtually any type of forest ecosystem in temperate, boreal and tropical areas (Gilbertson & Ryvarden, 1987). Trametes versicolor (“turkey tails”) is among the most common and widespread species of mushroom-forming fungi and its whole genome is currently being sequenced (http://www.jgi.doe.gov/). While the concept of Trametes just outlined is widely accepted (e.g., Ryvarden 1991, Ryvarden & Gilbertson 1994, Bernicchia 2005) it is not the only one that has been proposed. Corner (1989) had a much wider generic concept that synonymized under Trametes 15 other genera of Polyporales with trimitic hyphal systems, including taxa with stipitate basidiocarps (e.g. Microporus) and genera causing a brown-rot type of decay (Daedalea, Fomitopsis). Ryvarden (1991) defined the Trametes-group of the family Polyporaceae Corda by the combination of trimitic hyphal system and white-rot type of decay and there he included all the genera synonymized by Corner (1989) with the exception of the brown-rot Daedalea and Fomitopsis and the stipitate Lignosus. Ryvarden also included some genera not considered by Corner like Cryptoporus, Elmerina and Fomitella in the Trametes-group of the Polyporaceae.
Recent molecular studies have helped to clarify the higher-level and generic relationships of polyporoid fungi, and some of the genera considered as part of the Trametes-group have been placed outside the Polyporales: Trichaptum in the Hymenochaetales (Hibbett & Donoghue, 1995; Binder & al., 2005; Larsson & al., 2006) and Elmerina in the Tremellales (Larsson & al. 2004). Other genera in the Trametes-group or Trametes sensu Corner (1989) belong in the Polyporales but are not closely related to Trametes: Cerrena belongs in the phlebioid clade (Ko & Jung, 1999a; Lee & Lim 2010) and the brown-rot genera Daedalea and Fomitopsis are placed in the antrodia clade (Ko & Jung, 1999b; Hibbett & Donoghue, 2001; Binder & al., 2005). Based on the studies just mentioned, the articles by Tomšovský & al. (2006) and Sotome & al. (2008), and the results presented here the remainder genera of the Trametes-group belong in the core polyporoid clade.
The most inclusive phylogenetic studies with a focus on Trametes are those of Tomšovský & al. (2006), who studied ITS and nLSU sequences in eleven isolates representing seven European species, and Ko & Jung (1999b), who studied mtSSU sequences in seven isolates of Trametes. Collectively, these studies support the placement of most Trametes species in the core polyporoid clade, but also suggested that Trametes is paraphyletic or polyphyletic, with Lenzites and Pycnoporus species intermixed with Trametes species.
Molecular data also showed that some species classified in Trametes by Corner (1989), Gilbertson & Ryvarden (1987) and Ryvarden & Gilbertson (1994) do not belong in the genus: (i) Trametes trogii is more closely related to Coriolopsis gallica than to the other species of Trametes (Ko & Jung, 1999b; Tomšovský & al., 2006); (ii) Trametes consors belongs in the genus Cerrena (Ko & Jung, 1999a); (iii) Tomšovský (2008) showed that Trametes cervina is probably related to Ceriporiopsis and has been transferred to the new genus Trametopsis (Tomšovský, 2008).
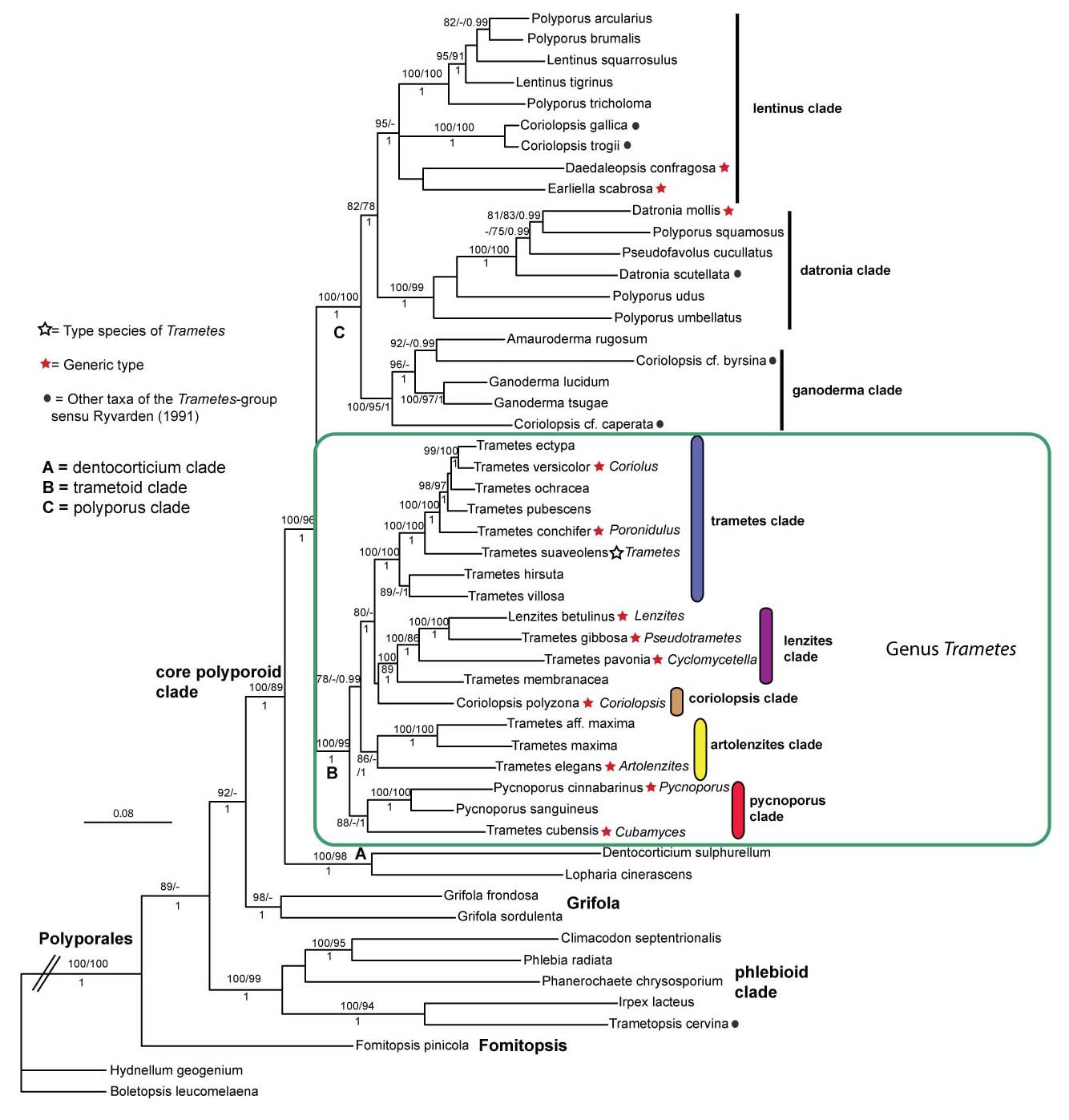
Discussion
These are the phylogenetic relationships of the Trametoid Polyporales infereed from nLSU, ITS, RPB1, RPB2 and TEF1 data.
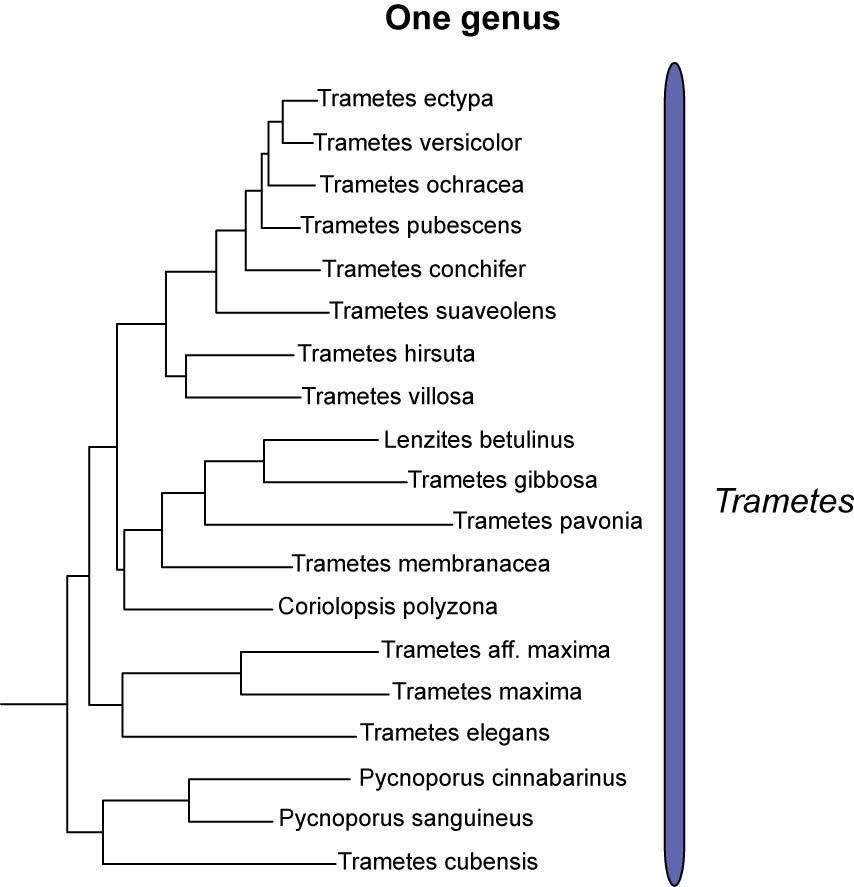
We conclude that all members of the trametoid clade should be classified under one single generic name (Trametes). The morphological concept of Trametes of Gilbertson & Ryvarden (1987) would only have to be slightly expanded to allow species with orange-red basidiocarps (Pycnoporus) and/or colored hyphae and context (Pycnoporus, C. polyzona) to be included in Trametes. Ascription of unsampled and newly described taxa to this “new” Trametes will be easier than in any of the alternative taxonomic scenarios considered here and not more complicated than with the current morphological concept of Trametes.
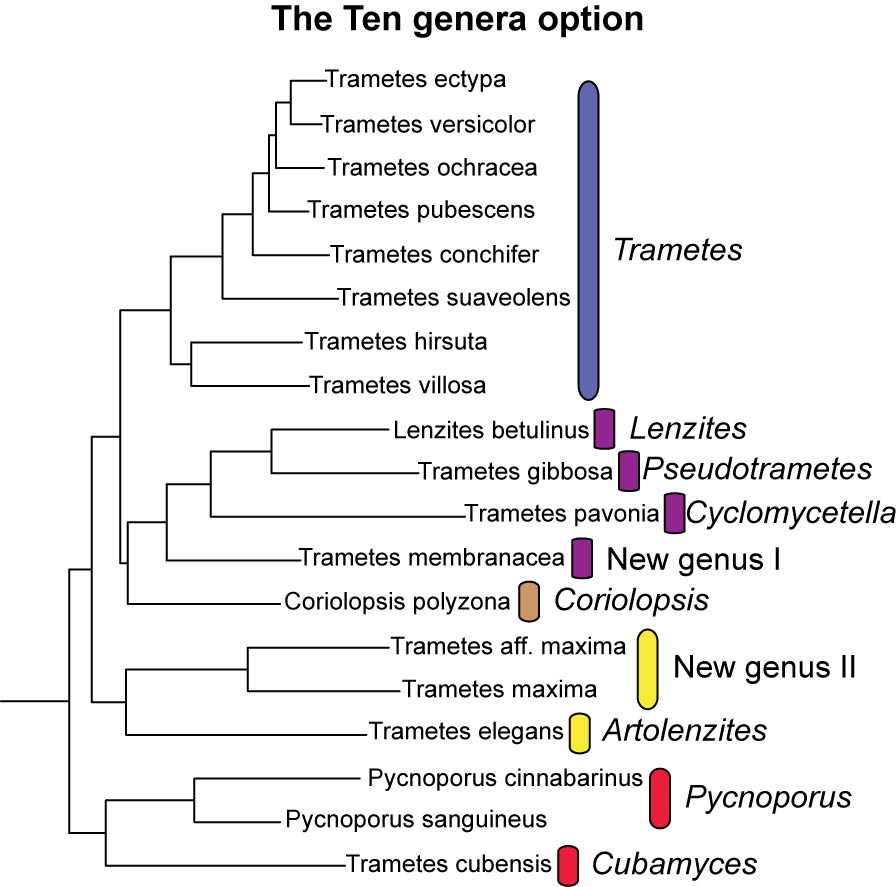
Alternatives to using the name Trametes for all members of the trametoid clade were also considered.
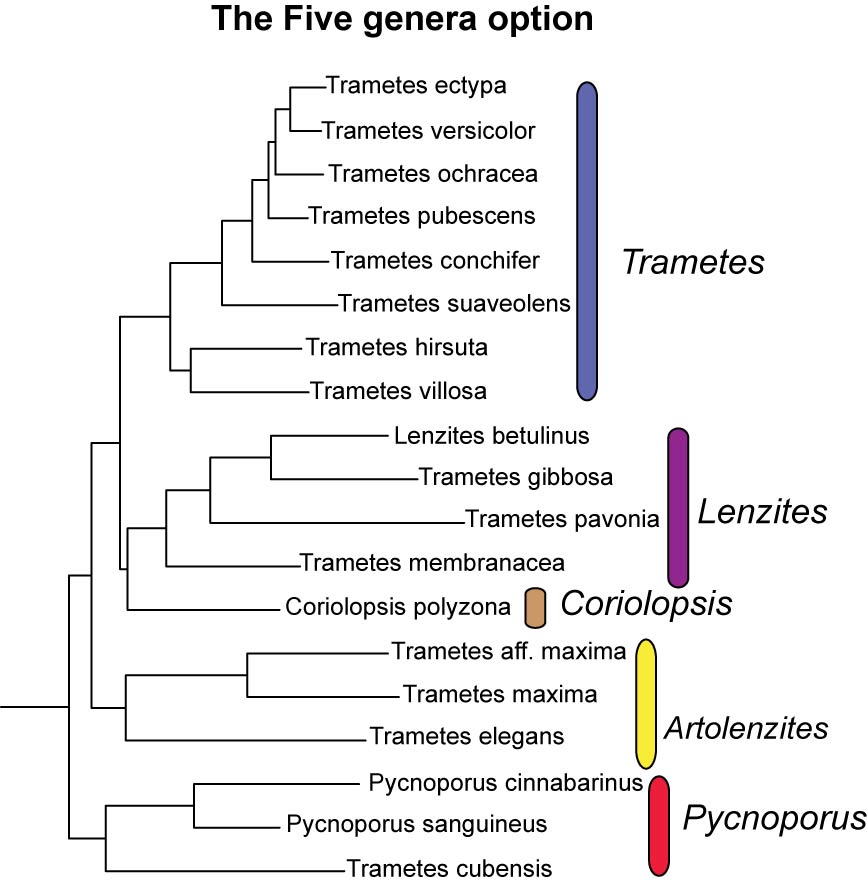
Five-genera option: recognize the five well-supported lineages recovered in the five-marker dataset analysis at the generic level: Trametes, Lenzites, Coriolopsis, Artolenzites and Pycnoporus. If this option is adopted it would be necessary to transfer some species currently placed in Trametes to Lenzites (T. gibbosa, T. pavonia, T. membranacea) or Pycnoporus (T. cubensis) and to “resurrect” the name Artolenzites to accommodate T. elegans and the T. maxima group. Reasons not to adopt this option can be readily recognized: (i) These five genera would be indistinguishable from each other based on morphology, ecology and/or biogeographical distribution; (ii) Given the homoplasious nature of some important characters used in the generic distinction of this group (e.g. lamellate vs. poroid hymenophore, presence of “sword-like” binding hyphae, nature and distribution of the pigments in the basidiocarps) it is not possible to determine, based on morphology alone, the taxonomic position of species not sampled here; (iii) It will not be possible to ascribe newly described species to any of these five genera without generating molecular data and even then if only rDNA is available (e.g. T. pocas, T. ljubarskii, L. warnieri) it may not be enough for a confident generic classification.
Ten-genera option: to recognize Trametes, Coriolopsis (only C. polyzona), Lenzites and Pycnoporus as defined by Gilbertson & Ryvarden (1987), without transferring any species into them, and adopting new generic names for the taxa that fall outside these genera. The generic names Pseudotrametes, Cyclomycetella, Artolenzites and Cubamyces would accommodate only their respective type species and two new genera would have to be created for T. membranacea and the T. maxima group. With the current sampling a total of ten genera would have to be accepted for taxa in the trametoid clade. However the same problems just outlined for the five genera option will be even more serious with this option and very likely the number of genera in the trametoid clade would increase as new taxa are sampled for molecular analyses.